Особенности векторных конструкцийСтраница 1

В зависимости от поставленной задачи используются два типа векторов: замещающий и вставочный. Первый тип векторов позволяет заменить участок гена мишени, в то время как второй интегрирует в изучаемую последовательность. Строение обоих типов векторов одинаково, кроме ориентации фланкирующих последовательностей (рис. 2). Наиболее часто используются замещающие вектора.
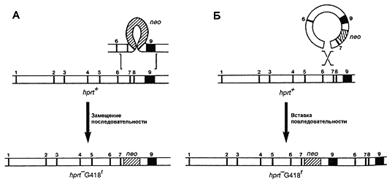
Рис. 2. Два типа векторов – замещающий (А) и вставочный (Б) – и их механизмы интеграции в геном [5].
Вектор для трансформации несет клонированную последовательность изучаемого гена, с внесенными в нее необходимыми изменениями. Это может быть: внесение стоп-кодона, приводящее к синтезу короткого неактивного пептида; делеция одного или нескольких экзонов; делеция промоторной области; вставка, приводящая к нарушению нормального функционирования гена и любые другие изменения, приводящие к отсутствию функционального продукта изучаемого гена или значительно снижающие его активность. Также в эту последовательность вносится положительный селективный маркер (МПС), которым является ген neo. Продукт этого гена дает несущим его клеткам устойчивость к антибиотикам неомицину и канамицину. Модифицированная последовательность должна быть фланкирована неизмененными участками, по которым будет проходить рекомбинация. Эффективность рекомбинации зависит от длины фланкирующих последовательностей [22], что в свою очередь зависит от возможностей вектора (рис. 3). При длине гомологичного плеча около 5 тыс. п.н. процент рекомбинации составляет 0,001.
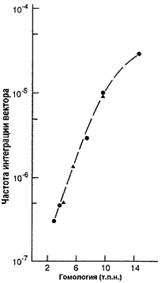
Рис. 3. Зависимость частоты интеграции вектора от длины гомологичных плеч [22].
В качестве вектора можно использовать бактериальные искусственные хромосомы (BAC), со вставками фрагментов генома мыши. В этом случае размер одного плеча может составлять до 150 тыс. п.н., а размер делеции до 25 тыс. п.н. Наилучший процент рекомбинации (8,3%) получен авторами с использованием длины плеча 110 тыс. п.н. [26].
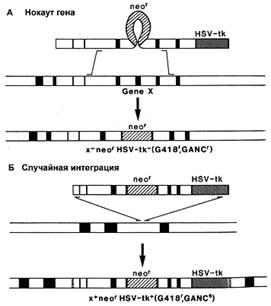
Существует вероятность, что рекомбинация пройдет не по исследуемым нами участкам генома, а в любой другой сходной области. При этом ген neo (МПС) сохранится, и в отобранном пуле ЭСК будут присутствовать рекомбинантные клетки, не несущие необходимых изменений. Эта проблема решается внесением в векторную конструкцию маркера отрицательной селекции (МОС). Им может служить ген тимидин-киназы простого вируса герпеса (HSV-tk) или ген дифтерийного токсина А (DT-A), продукты которых убивают эукариотические клетки. Положение МОС с наружной стороны гомологичного плеча вектора позволяет элиминировать его после прохождения гомологичной рекомбинации (рис. 4). В случае же негомологичной рекомбинации, МОС оказывается интегрированным в геном трансформированной клетки, что приводит к ее элиминации. Наличие двух маркеров селекции (положительного и отрицательного) позволяет быстро и эффективно проводить отбор нужных трансформантов [23,19].
Другие статьи:
Разложение трудноразлагаемых веществ
Плесневые грибы в качестве источника углерода могут использовать такие трудноразлагаемые вещества, как целлюлоза, крахмал, лигнин, пектиновые вещества, нефть, пестициды.
Разложение целлюлозы.
Основными источниками целлюлозы для грибов в ...
Способы разрушения тканей и клеток
Для разрушения клеток чаще всего применяют физические методы. Большинство животных клеток разрушается сравнительно легко, однако при разрушении растительных и бактериальных клеток зачастую приходится сталкиваться со значительными трудност ...
Гнездо пчелиной семьи
Гнездо пчёлы строят сами из воска. Оно состоит из сотов, и служит для пребывания взрослых пчел, выращивание расплода и складывания кормовых запасов. Каждый сот состоит из ячеек, расположенных на общем основании в два слоя.
Ячейки, предна ...