Гликозилирование белков — одна из главных причин поздних осложнений сахарного диабета

Поздние осложнения сахарного диабета связаны прежде всего с повреждением кровеносных сосудов (диабетические ангиопатии). Основной механизм повреждения тканей — гликирование (гликозилирование) белков — не ферментативная реакция глюкозы со свободными аминогруппами белковой молекулы (Лиз, Арг, N-концевая аминокислота):
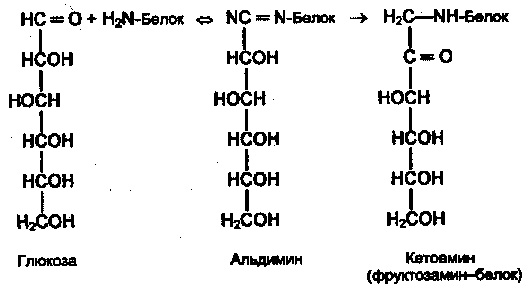
Вначале образуется нестабильная альдиминовая группировка, которая может превращаться в ряд других, более стабильных соединений («ранние продукты гликозилирования»). Понятно, что функции белка могут быть нарушены в результате изменения заряда белковой молекулы, ее конформации или блокирования активного центра. Гликозилирование — медленная реакция, в тканях здоровых людей обнаруживаются лишь небольшие количества гликозилированных белков. При гипергликемии реакция существенно ускоряется. Например, у больных диабетом в состоянии гипергликемии содержание одного из гликозилированных гемоглобинов — HbAlc — в течение 2—3 нед увеличивается в 2—3 раза. Степень гликозилирования разных белков неодинакова; в основном она зависит от скорости обновления данного белка. В медленно обменивающихся белках накапливается больше модифицированных аминогрупп. Кроме того, в таких белках происходят дальнейшие изменения углеводных остатков: перестройка структуры, окислительные превращения, в результате которых образуются разнообразные «поздние продукты гликозилирования» (ППГ), часто коричневого цвета, флюоресцирующие, и некоторые из них обладают высокой реакционной активностью и способностью дополнительно повреждать белки, в том числе образовывать поперечные сшивки между молекулами белков. К медленно обменивающимся белкам относятся многие белки соединительно-тканных образований, межклеточного матрикса, базальных мембран. К тому же белки этих структур непосредственно контактируют с межклеточной жидкостью, в которой концентрация глюкозы такая же, как в крови (в клетках она обычно гораздо ниже в результате использования глюкозы в метаболических процессах). В этих структурах ППГ накапливаются с возрастом, накопление сильно ускоряется при сахарном диабете.
ППГ-белки могут гидролизоваться макрофагами (с участием ППГ-рецепторов) или межклеточными протеолитическими системами с образованием ППГ-пептидов, часто длиной около 30 аминокислотных остатков. ППГ-белки, особенно образующиеся в результате их гидролиза ППГ-пептиды, попадают и в кровоток. Концентрация ППГ-пептидов в крови резко повышается при почечной недостаточности разного происхождения, в том числе при диабетической нефропатии. Это связано с тем, что элиминация ППГ-пептидов происходит с участием почек: ППГ-пептиды фильтруются в клубочках, реабсорбируются клетками проксимальных канальцев и катаболизи-руются в лизосомах этих клеток.
В экспериментах на крысах показано, что введение ППГ-белков в кровь приводит к ковалентному связыванию этих белков с белками межклеточного матрикса во многих тканях и к появлению структурных и функциональных нарушений, сходных с теми, которые бывают при сахарном диабете.
ППГ проявляют многообразную биологическую активность: повышают проницаемость эндотелиальных клеток, соединяются с рецепторами макрофагов, эндотелиальных и мезангиальных клеток, активируют макрофаги к секреции цитокинов (рецепторным путем), подавляют образование NО и соответственно ингибируют расширение сосудов, усиливают окисление ЛНП. В крови больных диабетом обнаруживаются антитела к ППГ-пептидам.
Другие статьи:
Анатомия
рис. 4
Дыхательная система человека состоит из тканей и органов, обеспечивающих легочную вентиляцию и легочное дыхание. К воздухоносным путям относятся: нос, полость носа, носоглотка, гортань, трахея, бронхи и бронхиолы. Легкие состоят ...
Роль фитобентоса в формировании продуктивности. Общие
сведения о первичной продукции
Первичная продукция водоемов — результат жизнедеятельности населяющих его растительных организмов — существенно отличается от всех других видов биологической продукции тем, что представляет собой новообразование органических веществ из ми ...
B6SJL-Tg(SOD1-G93A)dl1Gur/J
Окраска шерсти: разная: белая, коричневая, чёрная.
Происхождение: линия создана в лаборатории Mark E. Gurney при Северозападном университете США (Northwestern University, USA).
Метод модификации: трансгеноз. Трансгенные мыши G93A, экспр ...