Связывание липидов с внутримембранными белками
в бислоеСтраница 1

Этой теме были посвящены многочисленные исследования, в которых использовались разнообразные подходы. По существу во всех этих работах ставилась задача выяснить, есть ли у белков участки, специфически взаимодействующие с определенными липидами, и можно ли считать белково-липидные комплексы долгоживущими, т. е. обладают ли они временем жизни, сравнимым с временем оборота типичного фермента. Такие исследования проводились с помощью Н-ЯМР, ЭПР и флуоресцентных методов. Чтобы разобраться в полученных результатах, полезно рассмотреть термодинамику простой реакции обмена, где липид одного типа вытесняется с места связывания на белке другим липидом:
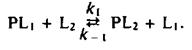
Константа равновесия этой реакции равна
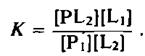
Она является относительной константой связывания липидов Li и L2 с данным участком белковой молекулы, причем
![]()
Если липиды двух типов присутствуют в мембране в одинаковой концентрации и сродство их к белку одинаково, то К = 1. Рассмотрим гипотетический случай, когда L2 является минорным липидным компонентом и составляет только 5°/о от общего количества липидов. Тогда
![]()
Предположим, что L2 предпочтительно связывается с неким участком белковой молекулы, так что в равновесии 90% участков занято L.2. Тогда
![]()
Следовательно, К = 171, что соответствует величине ДС° = = -3,1 ккал/моль. Столь небольшого различия в свободной энергии связывания оказывается достаточно для существенного смещения распределения липидов, связанных с белком, от равновесного состояния. Это обусловлено тем, что соотношение эффективных концентраций конкурирующих липидов в мембране относительно мало: не превышает 100, а в большинстве случаев гораздо меньше. Другими словами, даже минорные липиды присутствуют в мембране в концентрации, составляющей не менее 1% от концентрации основных липидов, с которыми они конкурируют за места связывания на белках.
Для оценки относительного сродства липидов к специфическим белкам используют два подхода. Они основаны на применении липидных аналогов, встроенных в фосфолипидные везикулы, которые содержат интересующий исследователя белок.
1. Спин-меченные фосфолипиды, соседствующие с мембранными белками, обладают ограниченной подвижностью. Это проявляется в уширении спектра ЭПР. У тех молекул ЭПР-зонда, которые соседствуют с белком, возможность движения с характерной частотой > 108 с" 1 существенно ограничена. Спектр ЭПР в этом случае может быть представлен в виде суммы двух компонент: компоненты с узкими спектральными линиями, соответствующей основной липидной фазе, и компоненты, отвечающей липидам с ограниченной подвижностью. Для того чтобы эта последняя была в спектре преобладающей, отношение белок/липид должно быть достаточно высоким. Осложняет картину то, что липиды могут попадать в «ловушки» из белковых агрегатов. Такие липиды тоже обладают ограниченной подвижностью, и им соответствует третья компонента в спектре ЭПР. Для выявления попавших в ловушку липидов при высоких концентрациях белка можно использовать липидные спиновые зонды, ковалентно связанные с поверхностью белка.
Другие статьи:
Основные стадии развития и простейшие методы исследования бактериофагов
Несмотря на чрезвычайное разнообразие бактериофагов, процедуры их исследования, за редким исключением, применимы к большинству фагов. В заводской лаборатории можно провести предварительное изучение фага с применением относительно простых ...
IL-8
Интерлейкин-8 (ультрачувствит.) ...
Профилактика нарушения опорно-двигательного
аппарата и детского травматизма
Правильная осанка-красота и стройность, свидетельство хорошего здоровья и высокой активности человека. Нарушение осанки разнообразны: сутуловатость, круглая спина, скомотическая осанка, лордотическая осанка.
Количество детей с нарушением ...