Происхождение жизниСтраница 3

Предлагаемая гипотетическая схема содержит два существенных момента, кажущихся принципиальными.
Во-первых, постулируется, что абиогенно синтезируемые олигорибонуклеотиды активно рекомбинировали посредством механизма спонтанной неэнзиматической трансэстерификации [18], приводя к образованию удлиненных цепей РНК и давая начало их многообразию. Именно этим путем в популяции олигонуклеотидов и полинуклеотидов и могли появиться как каталитически активные виды РНК (рибозимы), так и другие виды РНК со специализированными функциями (см. рис. 7). Более того, неэнзиматическая рекомбинация олигонуклеотидов, комплементарно связывающихся с полинуклеотидной матрицей, могла обеспечить сшивание (сплайсинг) фрагментов, комплементарных этой матрице, в единую цепь. Именно таким способом, а не катализируемой полимеризацией мононуклеотидов, могло осуществляться первичные копирование (размножение) РНК. Разумеется, если появлялись рибозимы, обладавшие полимеразной активностью [20], то эффективность (точность, скорость и продуктивность) копирования на комплементарной. матрице должна была значительно возрастать.
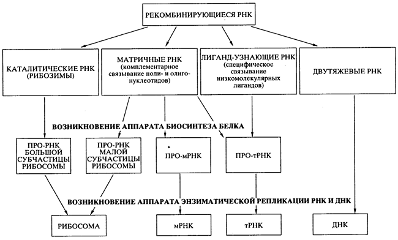
Рис. 7.
Схема эволюции и специализации молекул РНК в процессе перехода от древнего мира РНК к современному миру генетически детерминированного биосинтеза белков
Второй принципиальный момент в моей версии состоит в том, что первичный аппарат биосинтеза белка возник на базе нескольких видов специализированных РНК до появления аппарата энзиматической (полимеразной) репликации генетического материала - РНК и ДНК. Этот первичный аппарат включал каталитически активную прорибосомную РНК, обладавшую пептидил-трансферазной активностью; набор про-тРНК, специфически связывающих аминокислоты или короткие пептиды; другую прорибосомную РНК, способную взаимодействовать одновременно с каталитической прорибосомной РНК, про-мРНК и про-тРНК (см. рис. 7). Такая система уже могла синтезировать полипептидные цепи за счет катализируемой ею реакции транспептидации. Среди прочих каталитически активных белков - первичных ферментов (энзимов) - появились и белки, катализирующие полимеризацию нуклеотидов - репликазы, или НК-полимеразы.
Впрочем, возможно, что гипотеза о древнем мире РНК как предшественнике современного живого мира так и не сможет получить достаточного обоснования для преодоления основной трудности - научно правдоподобного описания механизма перехода от РНК и ее репликации к биосинтезу белка. Имеется привлекательная и детально продуманная альтернативная гипотеза А.Д. Альтштейна (Институт биологии гена РАН), в которой постулируется, что репликация генетического материала и его трансляция - синтез белка - возникали и эволюционировали одновременно и сопряженно, начиная с взаимодействия абиогенно синтезирующихся олигонуклеотидов и аминоацил-нуклеотидилатов - смешанных ангидридов аминокислот и нуклеотидов [22]. Но это уже следующая сказка . ("И Шахразаду застигло утро, и она прекратила дозволенные речи".
Другие статьи:
Кожа, как орган осязания
Как мы говорили ранее чувствительные нервные волокна, позволяющие человеку осязать, пронизывают кожу. Что же представляет собой сама кожа?
Кожа является самым большим органом человеческого тела. Общая площадь кожи взрослого составляет ок ...
Гомель как крупный промышленный центр
Гомель - второй по величине город Беларуси, важнейший промышленный и культурный центр Белорусского Полесья. Впервые упоминается в летописи в 1142г. Расположен на р. Сож. Здесь сосредоточены крупные предприятия машиностроения, легкой и пищ ...
Класс водохранилища. Группа типов искусственные пруды и
запруды
В пруды и водохранилища использовались главным образом для орошения и обводнения сельскохозяйственных угодий, рекреации и рыборазведения.
По количеству прудов и водохранилищ первое место занимает бассейн реки Бол.Алим. Здесь емкостью иск ...