Свойства ДНК как
вещества наследственности и изменчивости. Самовоспроизведение
наследственного материала. Репликация ДНКСтраница 2

Роль затравки для синтеза полинуклеотидных цепей ДНК в ходе репликации выполняют короткие последовательности РНК, образуемые при участии фермента РНК-праймазы (рис.11). Указанная особенность ДНК-полимеразы означает, что матрицей при репликации может служить лишь цепь ДНК, несущая спаренную с ней затравку, которая имеет свободный 3'-ОН-конец.
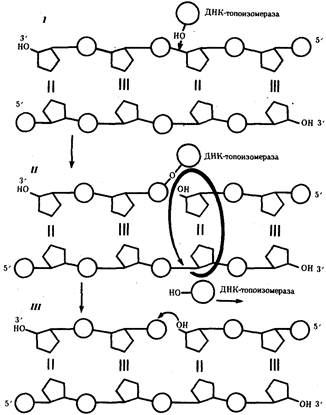
Рис.9. Разрыв одной из цепей ДНК с помощью фермента ДНК-топоизомеразы: I - ДНК-топоизомераза образует ковалентаую связь с одной из фосфатных групп ДНК (верхняя цепь); II - в результате разрыва фосфодиэфирной связи в одной полинуклеотидной цепи вокруг соответствующей ей связи другой цепи осуществляется вращение, которое снимает напряжение, вызванное расхождением двух цепей ДНК в области репликационной вилки; III - после снятия напряжения в спирали ДНК происходит спонтанное отделение ДНК-топоизомеразы и восстановление фосфодиэфирной связи в цепи ДНК
Способность ДНК-полимеразы осуществлять сборку полинуклеотида в направлении от 5' - к 3' - концу при антипараллельном соединении двух цепей ДНК означает, что процесс репликации должен протекать на них по-разному. Действительно, если на одной из матриц (3' → 5') сборка новой цепи происходит непрерывно от 5' - к 3'-концу и она постепенно удлиняется на 3'-конце, то другая цепь, синтезируемая на матрице (5' → 3'), должна была бы расти от 3' - к 5'-концу. Это противоречит направлению действия фермента ДНК-полимеразы.
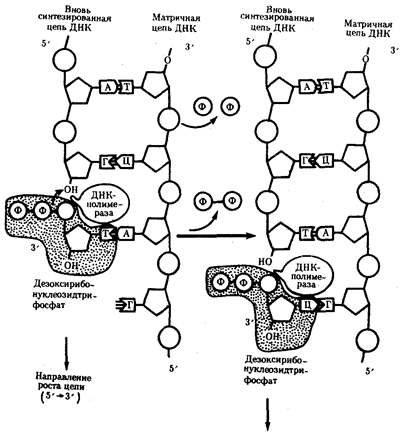
Рис.10. Присоединение очередного нуклеотида к дочерней цепи ДНК, синтезируемой при участии ДНК-полимеразы: ФФ-пирофосфат
В настоящее время установлено, что синтез второй цепи ДНК осуществляется короткими фрагментами (фрагменты Оказаки) также в направлении от 5' - к 3'-концу (по типу шитья "назад иголкой"). У прокариот фрагменты Оказаки содержат от 1000 до 2000 нуклеотидов, у эукариот они значительно короче (от 100 до 200 нуклеотидов). Синтезу каждого такого фрагмента предшествует образование РНК-затравки длиной около 10 нуклеотидов. Вновь образованный фрагмент с помощью фермента ДНК-лигазы соединяется с предшествующим фрагментом после удаления его РНК-затравки (рис.12, А).
В связи с указанными особенностями репликационная вилка является асимметричной. Из двух синтезируемых дочерних цепей одна строится непрерывно, ее синтез идет быстрее и эту цепь называют лидирующей. Синтез другой цепи идет медленнее, так как она собирается из отдельных фрагментов, требующих образования, а затем удаления РНК-затравки. Поэтому такую цепь называют запаздывающей (отстающей). Хотя отдельные фрагменты образуются в направлении 5' → 3', в целом эта цепь растет в направлении 3' → 5' (рис.3.12, А). В виду того, что от локуса ori как правило начинаются две репликационные вилки, идущие в противоположных направлениях, синтез лидирующих цепей в них идет на разных цепях материнской ДНК (рис 12, Б). Конечным результатом процесса репликации является образование двух молекул ДНК, нуклеотидная последовательность которых идентична таковой в материнской двойной спирали ДНК.
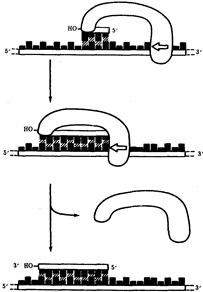
Рис.11. Схема реакции синтеза короткой РНК-затравки, катализируемой РНК-праймазой
Рассмотренная последовательность событий, происходящих в ходе репликативного синтеза, предполагает участие целой системы ферментов: геликазы, топоизомеразы, дестабилизирующих белков, ДНК-полимеразы и других, совместно действующих в области репликационной вилки (рис 13).
Репликация ДНК у про- и эукариот в основных чертах протекает сходно, однако, скорость синтеза у эукариот (около 100 нуклеотидов/с) на порядок ниже, чем у прокариот (1000 нуклеотидов/с). Причиной этого может быть образование ДНК эукариот достаточно прочных соединений с белками, что затрудняет ее деспирализацию, необходимую для осуществления репликативного синтеза.
Фрагмент ДНК от точки начала репликации до точки ее окончания образует единицу репликации - репликон. Однажды начавшись в точке начала (локус on), репликация продолжается до тех пор, пока весь репликон не будет дуплицирован. Кольцевые молекулы ДНК прокариотических клеток имеют один локус on и представляют собой целиком отдельные репликоны. Эукариотические хромосомы содержат большое число репликонов. В связи с этим удвоение молекулы ДНК, расположенной вдоль эукариотической хромосомы, начинается в нескольких точках. В разных репликонах удвоение может идти в разное время или одновременно.
Другие статьи:
Результаты исследования уровня гамма-интерферона в сыворотке крови у больных
с хроническим рецидивирующим фурункулезом
Результаты исследования уровня гамма-интерферона в сыворотке крови у больных с хроническим рецидивирующим фурункулезом приведены на рис. 9 и табл. 1 (Приложение 1).
Полученные результаты показали‚ что изменения уровня ИФНγ как у бол ...
Методы исследования городской авифауны
Разными авторами используются различные методы исследования птиц в условиях города. Ввиду неоднородности городских местообитаний обычно используют даже несколько методов учета. На застроенных территориях птиц обычно подсчитывают на трансе ...
Первые шаги климатологии.
Путешественники и мореплаватели древности уже весьма давно обратили внимание на различие климатов тех или других стран, которые им довелось посетить. Климатология, таким образом, в течение веков шла рука об руку с географией, будучи ее не ...