Конфигурация и упаковка ацильных цепей в бислое

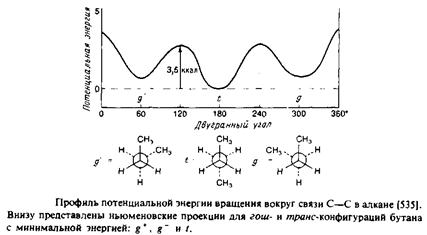
Рассмотрим сначала насыщенные углеводородные цепи. В них возможно свободное вращение вокруг каждой С—С-связи, характеризующееся энергетическим минимумом, особенно четким в случае ньюменовской проекции. Наиболее стабильна транс-конфигурация, при этом высота энергетического барьера для перехода через заслоненную конфигурацию в гош-форму составляет по оценкам 3,5 ккал/моль. В полностью m/wwc-конфигурации цепь максимально вытянута и не меняет своего направления, тогда как в гош-конформации ее направление меняется. Последовательность гош-транс-гош для трех смежных С—С-связей приводит к появлению в цепи излома, в результате чего участки цепи выше и ниже места излома оказываются значительно смещенными друг относительно друга. /ош-конфигурация в зависимости от направления вращения при переходе от Q к С4 обозначается как g+ или g". Кинки типа g + tg~ или g~ tg+ приводят к минимальному сдвигу цепи. Почти все двойные связи в мембранных липидах находятся в J/ис-конфигурации. Как и в случае гош-конфигурации, это приводит к изменению общего направления цепи. Наличие в углеводородных цепях кинков, двойных ^ыс-связей,
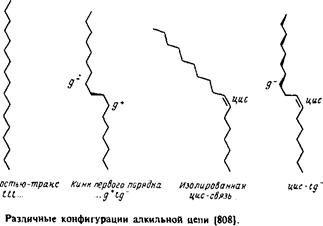
циклопропановых групп и других особенностей приводит к увеличению площади поперечного сечения цепи; это может иметь важные последствия для упаковки липидов в бислое. При этом стерические требования к упаковке углеводородных цепей и полярных головок такие же, как и в липидных крастал-лах. Эти принципы будут обсуждаться в разд. 2.3 при анализе формы мицелл.
Многие методы, включая дифракцию рентгеновских лучей и нейтронов, спектроскопию КР и ИК-спектрометрию, указывают, что в фазе геля насыщенные углеводородные цепи фосфолипидов находятся преимущественно в полностью-гпрднс-конфигурации. Минимальная площадь поперечного сечения молекулы диа-цильного фосфолипида равна около 38 A2. Примерно такую площадь занимает полярная головка фосфатидилэтанола-мина, поэтому насыщенные фосфатидилэтаноламины в гелевой фазе упаковываются так, что ацильные цепи располагаются перпендикулярно плоскости бислоя, как и в липидных кристаллах. В случае же кристаллов фосфатидилхолина минимальная площадь, приходящаяся на одну полярную головку, составляет примерно 50 А2. Поэтому дипальмитоилфосфатидилхолин в гелевой фазе не может упаковываться так, как фосфатидилэтаноламин. В этом случае ацильные цепи дипальмитоилфосфатидилхолина отклоняются на 30° от нормали к бислою, благодаря чему их поперечное сечение увеличивается и достигается соответствие размеру полярной головки. При этом углеводородные цепи сохраняют полнос-тью-транс-конфигурацию. В жидкокристаллической фазе появление в цепи гош-конформеров увеличивает эффективное поперечное сече-
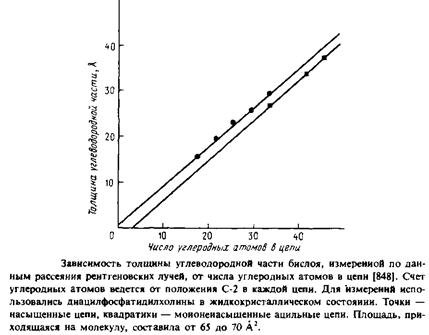
ние цепей по меньшей мере до 50 А2 в расчете на молекулу диациль-ного фосфолипида, а в водных дисперсиях эффективная площадь, приходящаяся на молекулу фосфолипида, составляет обычно 60 — 70 А2. Следовательно, в жидкокристаллической фазе углеводородные цепи не наклонены к плоскости бислоя, поскольку в этих условиях.полярные головки липидных молекул достаточно удалены друг от друга, и чтобы заполнить пространство между соседними головками и перекинуть между ними мостики, требуются вода и другие полярные молекулы. Судя по данным 2Н-ЯМР, толщина углеводородной области дипальмитоилфосфатидилхолинового бислоя в жидкокристаллическом состоянии составляет 35, а не 45 А, как следовало ожидать, если бы цепи находились в полностью-/я/*7«с-конфиругации и были ориентированы вдоль нормали к би-слою. Толщина бислоя уменьшается за счет наличия к цепях гош-конформеров, приводящих к разупорядоченности цепей, причем сами цепи в целом растянуты и расположены перпендикулярно поверхности бислоя, а не скручены в спираль. На рис. 2.12 приведена линейная зависимость толщины жидкокристаллического бислоя в диацилфосфатидилхолиновых везикулах от длины ацильных цепей, построенная по данным рассеяния рентгеновских лучей.
Другие статьи:
Снятие промеров
Сразу после отстрела животного, предназначенного для изготовления чучела, до наступления трупного окоченения, рулеткой или мерной лентой необходимо произвести следующие измерения:
Когда закончено снятие промеров, желательно сделать конту ...
Главнвые направления эволюции семейства Полиноид
Положение в общей системе полихет чешуйчатых червей и близких к ним семейств, объединенных нами в подотряд Aphroditiformia, уже было рассмотрено ранее. Здесь остановимся на филогенетических отношениях в наиболее богатом родами семействе P ...
Общие закономерности роста и развития школьников. Рост и развитие
В возрастной физиологии и школьной гигиене понятия рост и развитие более конкретны, чем в других науках. Рост - это изменения анатомических и морфологических показателей, т. е длины и веса тела. Он является одним из основных возрастно-пол ...