Происхождение эукариотической клеткиСтраница 1

Ископаемые останки клеток эукариотического типа обнаружены в породах, возраст которых не превышает 1,0–1,4 млрд. лет. Более позднее возникновение, а также сходство в общих чертах их основных биохимических процессов (самоудвоение ДНК, синтез белка на рибосомах) заставляют думать о том, что эукариотические клетки произошли от предка, имевшего прокариотическое строение.
Наиболее популярна в настоящее время симбиотическая гипотеза происхождения эукариотических клеток, согласно которой (рис. 3) основой, или клеткой-хозяином, в эволюции клетки эукариотического типа послужил анаэробный прокариот, способный лишь к амебоидному движению. Переход к аэробному дыханию связан с наличием в клетке митохондрии, которые произошли путем изменений симбионтов – аэробных бактерий, проникших в клетку-хозяина и сосуществовавших с ней.
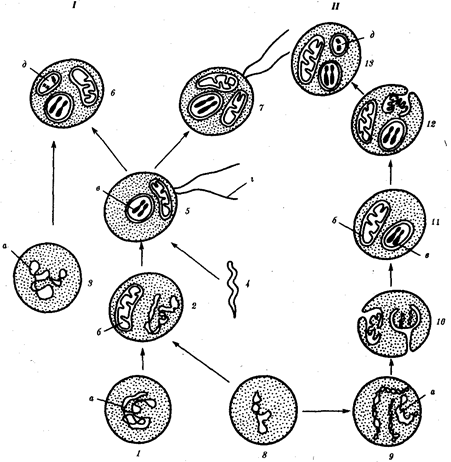
Рис. 3 Происхождение эукариотической клетки согласно симбиотической (I) и инвагинационной (II) гипотезам:
1 – анаэробный прокариот (клетка-хозяин), 2 – прокариоты, имеющие митохондрии, 3 – сине-зеленая водоросль (презумптивный хлоропласт), 4 – сиирохетообразная бактерия (презумптивный жгутик), 5 – примитивный эукариот со жгутиком, 6 – растительная клетка, 7 – животная клетка со жгутиком, 8 – аэробный прокариот (презумптивная митохондрия), 9 – аэробный прокариот (клетка-родоначальница согласно гипотезе II), 10 – инвагинации клеточной оболочки, давшие ядро и митохондрии, 11 – примитивный эукариот 12 – впячивание клеточной оболочки, давшее хлоропласт, 13 – растительная клетка; а–ДНК прокариотической клетки, б – митохондрия, в - ядро эукариотической клетки, г – жгутик, д – хлоропласт
Сходное происхождение предполагают для жгутиков, предками которых служили симбионты-бактерии, имевшие жгутик и напоминавшие современных спирохет. Приобретение клеткой жгутиков имело наряду с освоением активного способа движения важное следствие общего порядка. Предполагают, что базальные тельца, которыми снабжены жгутики, могли эволюционировать в центриоли в процессе возникновения механизма митоза.
Способность зеленых растений к фотосинтезу обусловлена присутствием в их клетках хлоропластов. Сторонники симбиотической гипотезы считают, что симбионтами клетки-хозяина, давшими начало хлоропластам, послужили прокариотические сине-зеленые водоросли.
Серьезным доводом в пользу симбиотического происхождения митохондрий, центриолей и хлоропластов является то, что перечисленные органеллы имеют собственную ДНК. Вместе с тем белки бациллин и тубулин, из которых состоят жгутики и реснички соответственно современных прокариот и эукариот, имеют различное строение. У бактерий не найдено также структур со свойственной жгутикам, ресничкам, базальным тельцам и центриолям эукариотических клеток комбинацией микротрубочек: «9 + 2» или «9 + 0».
Внутриклеточные мембраны гладкой и шероховатой цитоплазматической сети, пластинчатого комплекса, пузырьков и вакуолей рассматривают как производные наружной мембраны ядерной оболочки, которая способна образовывать впячивания.
Центральным и трудным для ответа является вопрос о происхождении ядра. Предполагают, что оно также могло образоваться из симбионта-прокариота. Увеличение количества ядерной ДНК, во много раз превышающее в современной эукариотической клетке ее количество в митохондрий или хлоропласте, происходило, по-видимому, постепенно путем перемещения групп генов из геномов симбионтов. Нельзя исключить, однако, что ядерный геном формировался путем наращивания генома клетки-хозяина (без участия симбионтов).
Согласно инвагинационной гипотезе, предковой формой эукариотической клетки был аэробный прокариот. Внутри такой клетки-хозяина находилось одновременно несколько геномов, первоначально прикреплявшихся к клеточной оболочке. Органеллы, имеющие ДНК, а также ядро, возникли путем впячивания и отшнуровывания участков оболочки с последующей функциональной специализацией в ядро, митохондрий, хлоропласты. В процессе дальнейшей эволюции произошло усложнение ядерного генома, появилась система цитоплазматических мембран.
Инвагинационная гипотеза хорошо объясняет наличие в оболочках ядра, митохондрий, хлоропластов, двух мембран. Однако она не может ответить на вопрос, почему биосинтез белка в хлоропластах и митохондриях в деталях соответствует таковому в современных прокариотических клетках, но отличается от биосинтеза белка в цитоплазме эукариотической клетки.
История показала, что эволюционные возможности клеток эукариотического типа несравнимо выше, чем прокариотического. Ведущая роль здесь принадлежит ядерному геному эукариот, который во много раз превосходит по размерам геном прокариот. Количество генов у бактерии и в клетке человека, например, соотносится как 1: (100–1000). Важные отличия заключаются в диплоидности эукариотических клеток благодаря наличию в ядрах двух комплектов генов, а также в многократном повторении некоторых генов. Это расширяет масштабы мутационной изменчивости без угрозы резкого снижения жизнеспособности, эволюционно значимым следствием чего является образование резерва наследственной изменчивости.
Другие статьи:
Словарь основных терминов и понятий. Животные и человек
Моллюски - тип беспозвоночных мягкотелых животных, большинство из которых имеют раковину различной формы. У голого слизня раковины нет, а у «корабельного червя» она сохранилась только на передней части головы. Тело моллюсков состоит из го ...
Отряд «блохи» (aphaniptera)
БЛОХИ — активные кровососы, паразитируют на многих животных, являются переносчиками возбудителей многих инфекционных болезней человека и животных (чума, туляремия, миксоматоз, эндемический блошиный тиф и др.).
БЛОХИ — переносчики возбуди ...
Физиология почек
В процессе жизнедеятельности в организме человека образуются значительные количества продуктов обмена, которые уже не используются клетками и должны быть удалены из организма. Кроме того, организм должен быть освобожден от токсичных и чуж ...