Инсулин и глюкагон как регуляторы депонирования и мобилизации гликогена и
жировСтраница 1

Инсулин участвует в регуляции таких клеточных процессов, как метаболизм, трансмембранный перенос ионов, аминокислот, глюкозы, синтез и распад белков. С влиянием на ядерные процессы — репликацию и транскрипцию – связано участие инсулина в регуляции клеточной пролиферации и дифференцировки, а также трансформации клеток. В патогенезе основных клинических проявлений сахарного диабета в наибольшей мере проявляется нарушение инсулиновой регуляции обмена глюкозы, жиров и аминокислот, связанного с энергетическим обменом. В результате согласованной работы разных органов и систем в организме поддерживается энергетический гомеостаз, под которым понимают соответствие между потребностью в энергии и обеспеченностью организма энергоносителями. Гомеостаз сохраняется даже при существенных изменениях в приеме пищи и энергетических затратах. Инсулин, а также тесно взаимодействующий с ним «контринсулярный» гормон глюкагон — главные регуляторы изменений метаболизма при смене состояний пищеварения и голодания (абсорбтивное и постабсорбтивное состояния). На пищеварение приходится 10—15 ч в сутки, а расход энергии происходит в течение всех 24 ч (с определенным снижением в часы ночного сна). Поэтому часть энергоносителей во время пищеварения складируется для использования в постабсорбтивном состоянии. Печень, жировая ткань и мышцы — главные органы, связанные с этими изменениями. Режим запасания включается после приема пищи и сменяется режимом мобилизации запасов после завершения пищеварения. Следовательно, у человека при обычном трехразовом питании смена режимов происходит трижды за сутки. Однако смена режимов выражена нечетко, поскольку в течение дня промежутки между приемами пищи небольшие (5 – 6 ч) и постабсорбтивный период едва успевает начаться (если вообще успевает), как наступает время очередного приема пищи. Типичным постабсорбтивным состоянием считают состояние утром до завтрака, после примерно десятичасового ночного перерыва в приеме пищи. Еще более наглядна модель ритма питания, которой придерживался великий немецкий философ Э. Кант: он принимал пищу один раз в сутки. За сутки исчерпываются запасы гликогена в организме, единственным источником глюкозы становится глюконеогенез, глюкоза используется преимущественно нервными клетками, в то время как почти все другие клетки получают энергию за счет окисления жирных кислот, а также кетоновых тел, образующихся в печени из жирных кислот. Такое состояние можно считать как постабсорбтивное или как кратковременное голодание. Эту модель (рис.1) мы и будем иметь в виду, рассматривая смену режимов обмена энергоносителей.
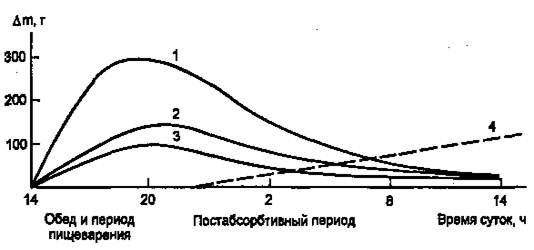
Рисунок 1. Изменение количества энергоносителей в организме человека (в тканях, не в желудке и кишечнике) в течение суток после однократного приема пищи. (1 — гликоген; 2 — жиры; 3 — аминокислоты/белки; 4 — изменение скорости глюконеогенеза, г/сут.)
Мышечная работа во время пищеварения замедляет процессы запасания, так как в мышцах непосредственно расходуется часть поступающих из кишечника продуктов переваривания. В постабсорбтивном состоянии мышечная работа стимулирует мобилизацию запасов, главным образом жиров. В регуляции изменений, связанных со сменой покоя и мышечной работы, важная роль принадлежит адреналину.
Потребление глюкозы клетками происходит при участии специальных белков-переносчиков (их называют также рецепторами глюкозы), образующих гидрофильные трансмембранные каналы. Существует два основных механизма переноса глюкозы: активный транспорт, зависящий от градиента концентраций ионов Na+, и облегченная диффузия. Соответственно есть два основных типа рецепторов глюкозы. Рецепторы, зависимые от концентрации ионов Na+, обнаруживаются только в почках и кишечнике, они обеспечивают реабсорбцию глюкозы из почечных канальцев и всасывание ее из просвета кишечника против градиента концентрации. Рецепторы облегченной диффузии (глюкозные транспортеры — ГЛЮТ) есть во всех тканях.
В тканях человека обнаружено пять разных ГЛЮТ:
ГЛЮТ-1 — в плаценте, мозге, почках, толстой кишке, в Р-клетках островков Лангерганса; меньше их в жировой ткани и мышцах;
ГЛЮТ-2 — преимущественно в печени, энтероцитах, в проксимальных тубулярных клетках почек (все эти клетки выделяют глюкозу в кровь); в Р-клетках панкреатических островков (островков Лангерганса). Возможно, участвует в стимуляции глюкозой секреции инсулина;
ГЛЮТ-3 во многих тканях, включая мозг, плаценту, почки;
ГЛЮТ-4 — единственный переносчик, регулируемый инсулином; содержится только в мышцах (скелетных и сердечной) и жировой ткани (инсулинзависимые ткани);
Все рецепторы могут находиться как в плазматической мембране клетки, так и в мембранных везикулах в цитоплазме. Количество рецепторов 1, 2, 3 и 5 в плазматической мембране изменяется в узких пределах и не зависит от концентрации инсулина. Напротив, ГЛЮТ-4 (и в гораздо меньшей степени ГЛЮТ-1) в отсутствие инсулина практически полностью находятся в цитозольных везикулах. Стимуляция клеток инсулином приводит к транслокации везикул к плазматической мембране и их слиянию, в результате чего рецепторы оказываются встроенными в плазматическую мембрану. Как показано в экспериментах с жировыми и мышечными клетками, скорость потребления глюкозы при этом увеличивается в 30—40 раз. При снижении концентрации инсулина в среде рецепторы вновь возвращаются в цитозоль.
Другие статьи:
Условия существования законов
Законы Менделя в их классической форме действуют при наличии определенных условий. К ним относятся:
1) гомозиготность исходных скрещиваемых форм;
2) образование гамет гибридов всех возможных типов в равных соотношениях (обеспечивается п ...
Обонятельные рецепторы
У позвоночных животных запахи выявляются структурой, содержащей примерно 100000 обонятельных рецепторных нейронов, аксоны которых проецируются через тонкий участок фронтального черепа (ситовидную пластинку) в обонятельную луковицуДлинные ...
Заглянем в историю
Теофраст (372–287 гг. до н.э.) определял ботанику как науку о растениях. Это и запечатлено в ее названии. Говоря о растениях, он несомненно имел в виду особей растений, а не виды, поскольку систематики растений в нашем понимании тогда про ...