Инсулин и глюкагон как регуляторы депонирования и мобилизации гликогена и
жировСтраница 2

Основными энергоносителями являются глюкоза и жирные кислоты. На рис. 2 представлены пути превращений глюкозы и жиров, а также белков и аминокислот.
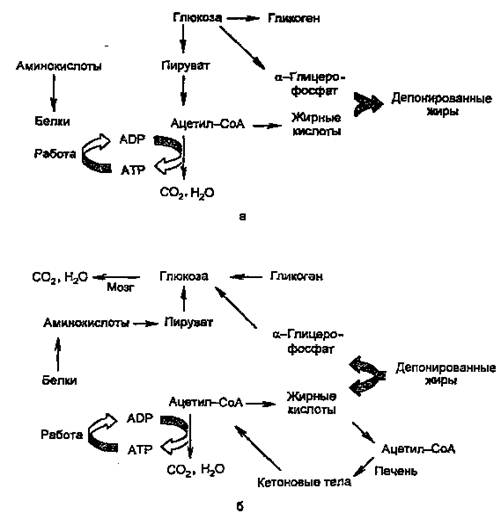
Рисунок 2. Изменение метаболизма основных энергоносителей при смене абсорбтивного (а) и постабсорбтивного (б) состояний.
Как видно из рисунка, при смене режимов многие процессы меняют направление на противоположное. За каждой из стрелок — серия реакций; ферменты, катализирующие ключевые реакции (лимитирующие скорость данной метаболической цепи), находятся под контролем многих регулирующих механизмов, включающих в качестве первого (внеклеточного) вестника сигнала главным образом инсулин и глюкагон, а также адреналин и кортизол.
Первичными сигналами для смены состояний являются изменение концентрации глюкозы в крови и вызванные этим реципрокные изменения концентраций инсулина и глюкагона. Регуляцию метаболизма инсулином и глюкагоном невозможно рассматривать по отдельности. В крови постоянно присутствуют оба гормона, однако изменяются их относительные концентрации. Действие каждого из них часто направлено на одни и те же конкретные мишени. Например, инсулин через путь Ras одновременно активирует гликогенсинтазу и ингибирует гликогенфосфорилазу, а глюкагон через сАМР-зависимые протеинкиназы одновременно ингибирует гликогенсинтазу и активирует гликогенфосфорилазу. Другой пример: инсулин сокращает не базальную скорость глюконеогенеза, а только скорость, стимулированную глюкагоном. На рис. 3 показаны некоторые другие мишени метаболических путей глюкозы в печени, общие для инсулина и глюкагона. Кроме того, инсулин снижает секрецию и самого глюкагона.
Глюкоза проникает в гепатоциты путем облегченной диффузии при участии ГЛЮТ-2, не зависимого от инсулина и имеющего высокую Км. В гепатоцитах глюкоза быстро превращается в глюкозо-6-фосфат глюкокиназой (гексокиназой IV), которая тоже имеет высокую Км (12 мМ) и не ингибируется продуктом реакции (в отличие от гексокиназ I, II и III). Далее глюкозо-6-фосфат может использоваться по трем направлениям: синтез гликогена, гликолиз, пентозофосфатный путь. Следует отметить, что ацетил-СоА, образующийся из глюкозы, используется для синтеза жирных кислот и жиров. Все эти пути стимулируются инсулином на пре- или посттрансляционном уровне.
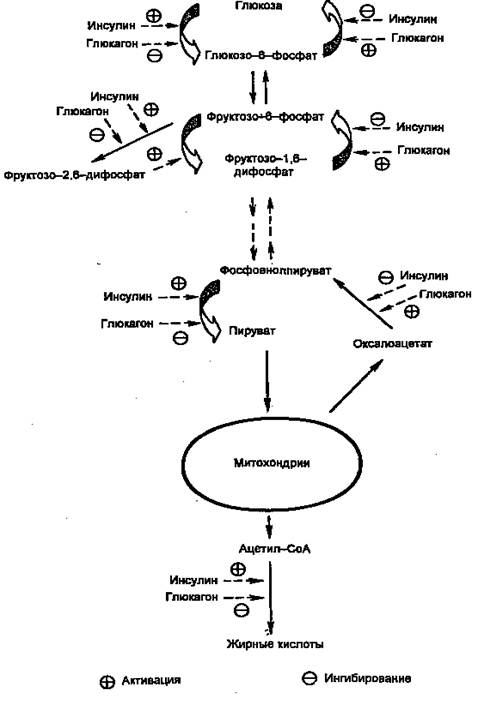
Рисунок 3. Действие инсулина и глюкагона на метаболизм глюкозы в печени.
Регуляция на претрансляционном уровне в свою очередь может быть двух типов: стимуляция транскрипции и повышение стабильности мРНК. В печени необратимые реакции гликолиза, а также синтез гликогена и синтез жиров стимулируются инсулином и подавляются глюкагоном. Наоборот, необратимые стадии глюконеогенеза подавляются инсулином и стимулируются глюкагоном. Подобная ситуация имеет место и в метаболизме жиров и аминокислот (белков): инсулин стимулирует их синтез, а глюкагон — мобилизацию. Поэтому направление метаболических процессов в сторону запасания или мобилизации зависит не столько от абсолютной концентрации гормона, сколько от отношения их концентраций ([инсулин]/[глюкагон], инсулин/глюкагоновый индекс). Для того, чтобы понять механизмы метаболических нарушений при инсулин-зависимом диабете, необходимо рассмотреть механизмы синтеза и секреции инсулина, так как нарушение этих процессов — одна из причин развития заболевания.
Другие статьи:
Возникновение теории эволюции и ее значение
Эволюционная биология, как и любая другая наука, прошла длинный и извилистый путь развития. Возникали и проверялись различные гипотезы. Большинство гипотез не выдерживало проверки фактами, и лишь немногие из них становились теориями, неиз ...
Механистическая картина мира
Согласно Ньютону, весь мир состоит «из твердых, весомых, непроницаемых, подвижных частиц». Эти «первичные частицы абсолютно тверды: они неизмеримо более тверды, чем тела, которые из них состоят, настолько тверды, что они никогда не изнаши ...
Какие типы взаимодействия являются короткодействующими? Привести примеры
систем, в которых действуют эти силы
Слабое взаимодействие менее известно за пределами узкого круга физиков и астрономов, но это нисколько не умаляет его значения. Достаточно сказать, что если бы его не было, погасли бы Солнце и другие звезды, ибо в реакциях, обеспечивающих ...