Синтез и секреция инсулинаСтраница 2

На рис. 5 показано изменение концентрации инсулина в крови человека после приема пищи. Одновременно со стимуляцией β-клеток к секреции инсулина происходит ингибирование секреции глюкагона из α-клеток панкреатических островков.
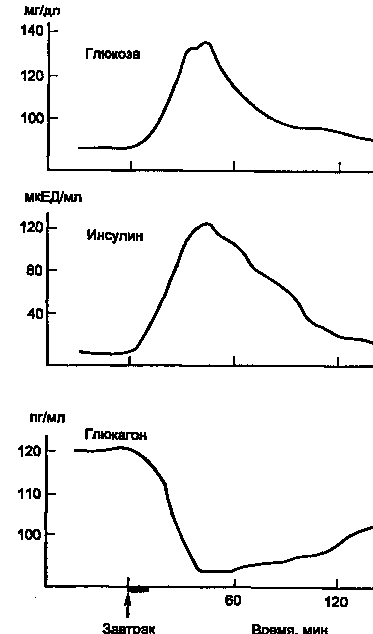
Рисунок 5. Изменение концентрации в крови глюкозы, инсулина и глюкагона после приема пищи (1 ЕД инсулина содержит 0,4081 мг белка инсулина).
Время полураспада инсулина в крови составляет 3—10 мин, а С-пептида — около 30 мин. Кровь при однократном прохождении через печень теряет до 60 % инсулина. В почках задерживается до 40 % инсулина, содержащегося в протекающей через почки крови, причем в клубочках инсулин фильтруется, а затем наряду с другими белками первичной мочи (альбумин, гемоглобин и др.) реабсорбируется и разрушается в клетках проксимальных канальцев нефрона.
Регуляция секреции инсулина зависит от глюкозосенсорной системы β-клеток, обеспечивающей пропорциональность между концентрацией глюкозы в крови и секрецией инсулина. Потребление глюкозы β-клетками происходит при участии ГЛЮТ-1 (основной переносчик глюкозы в β-клетках человека) и, возможно, ГЛЮТ-2. Эта ступень не является лимитирующей: концентрация глюкозы в клетке быстро уравнивается с концентрацией в крови. В β-клетках глюкоза превращается в глюкозо-6-фосфат глюкокиназой (гексокиназой IV, как и в глюкозосинтезирующих органах — печени, почках), имеющей высокую Км для глюкозы — 12 мМ (Км гексокиназ I, II и III — от 0,2 до 1,2 мМ). Вследствие этого скорость фосфорилирования глюкозы практически линейно зависит от ее концентрации в крови. Кроме того, глюкокиназа в Р-клетках — лимитирующее звено гликолиза. Поэтому глюкокиназа — вероятно, основной (но не единственный) элемент глюкозосенсорной системы β-клеток. Мутации глюкокиназы приводят к развитию одной из форм сахарного диабета — диабету I типа у взрослых (MODY).
Специфический ингибитор глюкокиназы манногептулоза подавляет стимуляцию глюкозой синтеза и секреции инсулина. Это указывает на то, что молекулы, непосредственно регулирующие синтез и секрецию инсулина, образуются в результате метаболизма глюкозы. Природа этих молекул неизвестна. Согласно имеющимся представлениям, роль такой молекулы может выполнять АТР (точнее, отношение ([ATP]/[ADP]). Гипотеза обосновывается тем, что секреция инсулина стимулируется только метаболизируемыми веществами — источниками энергии. Например, глюкоза и глицеральдегид стимулируют секрецию пропорционально скорости их метаболизма. Глицерин не метаболизируется в Р-клетках (низкая активность глицеролкиназы) и не стимулирует секрецию инсулина. Однако после обработки рекомбинантным аденовирусом, содержащим бактериальный ген глицеролкиназы, клетки приобретают способность отвечать на глицерин секрецией инсулина в такой же мере, как и на глюкозу.
Есть указание на участие в регуляции секреции инсулина не только гликолиза, но и митохондриальных процессов. В частности, существенное значение могут иметь анаплеротические (восполняющие, компенсирующие) реакции: пируват → оксалоацетат, глутамат → α-кетоглутарат Эти реакции увеличивают количество компонентов цитратного цикла, а следовательно, и его мощность. Стимулированная глюкозой секреция инсулина усиливается некоторыми аминокислотами, жирными кислотами, кетоновыми телами; таким образом, в стимуляции секреции участвует не только глюкоза, но все основные энергоносители. Следовательно, количество секретируемого инсулина пропорционально энергетической ценности потребляемой пищи. Окисление основных энергоносителей в цикле лимонной кислоты, усиленном анаплеротическими реакциями, может быстро привести к изменению отношений ATP/ADP и NADH/NAD+ в клетке. Изменение концентрации этих веществ в свою очередь приводит к появлению вторых вестников сигнала (возможно, ионов Са2+, сАМР, диацилглицерола, инозитол-3-фосфата), которые включают процесс экзоцитоза инсулиновых гранул.
Механизмы активации экзоцитоза остаются неясными. Ряд экспериментальных данных указывает на участие Са2+/кальмо-дулинзависимой протеинкиназы (СаМПК), а также полифункциональной СаМПК II, которая найдена в панкреатических островках крысы и активируется глюкозой.
Глюкокиназа — основной элемент глюкозосенсорного механизма Р-клеток; она имеется также и в β-клетках, а гликолиз ускоряется пропорционально внеклеточной концентрации глюкозы и в тех, и в других клетках. Между тем секреция гормона (инсулина и глюкагона соответственно) стимулируется глюкозой в β-клетках и подавляется в β-клетках. Возможно, это связано с тем, что в β-клетках в отличие от а-клеток очень высокая активность пируваткарбоксилазы (анаплеротический фермент), сравнимая с активностью в клетках, для которых характерен глюконеогенез (печень, почки). При этом наблюдается пропорциональность между увеличением концентрации цитрата и малата в клетках и секрецией инсулина. Можно думать, что какие-то метаболиты этих путей или связанная с ними активация пируватмалатного челночного механизма участвует в сопряжении стимула с секрецией инсулина.
Другие статьи:
Ч. Дарвин основатель теории биологической эволюции
Важным источником формирования естественно-научных основ психологии явилось эволюционное учение Чарльза Дарвина (1809-1882). В 1859 г. в свет выходит его книга "Происхождение видов путем естественного отбора", вероятно, самая зн ...
IFN-g
Интерферон-g ...
Приложения
Табл. Результаты работы.
№
Объект
Бактерии
Актиномицеты
Плесневые грибы
0
1
2
3
4
1
Руки грязные
8 колоний
-
7 колоний
2
Руки чистые
3 колонии
-
6 колоний
3
Зубной ...