Введение

Все биологические структуры по своей природе динамичны, и при рассмотрении их функций необходимо учитывать подвижность компонентов, из которых эти структуры состоят. Это относится к ферментам, полинуклеотидам и, конечно, к мембранам. В жидкостно-мозаичной модели, в центре которой находится представление о подвижности мембранных компонентов, мембрана рассматривается как некое липидное море, в котором свободно плавают глобулярные белки. За последние два десятилетия было опубликовано множество работ, посвященных количественным и качественным аспектам динамики мембранных компонентов. Во многих из них использовались спектральные методы и спектроскопия магнитного резонанса. В результате была создана весьма полезная физическая картина мембран, иллюстрирующая способы перемещения мембранных белков и липидов и их взаимодействия. Кроме того, были разработаны специальные методики, направленные на изучение динамических свойств мембран. Многое в этой области остается неясным, но уже определены основные направления будущих исследований.
Главным побудительным мотивом в изучении мембранной динамики служит ее связь с биологическими функциями мембран. Необходимым условием протекания одних ферментативных процессов является свободная диффузия мембраносвязанных компонентов в плоскости бислоя, другие же процессы могут осуществляться лишь при ограниченной подвижности мембранных компонентов. Между шероховатым и гладким эндоплазматическими ретикулумами, комплексом Гольджи и плазматической мембраной происходит быстрый обмен различными веществами, и тем не менее их состав и функции различаются. Чтобы понять суть этих и многих других биологических феноменов, необходимо прежде всего выяснить фундаментальные аспекты динамических свойств мембран. Поперечная асимметрия в распределении липидов, а возможно, и пассивная проницаемость бислоев очевидным образом связаны со скоростью трансмембранного флип-флоп-переноса липидов. Биогенез мембран зависит от скорости обмена липидов между различными мембранами. Скорость ферментативных реакций, протекающих с участием мембраносвязанных компонентов, зависит от скорости латеральной диффузии компонентов мембран. Наконец, липидно-белковые взаимодействия зависят от скорости, с которой происходит обмен липидами между ближайшим окружением белков и остальным объемом мембраны.
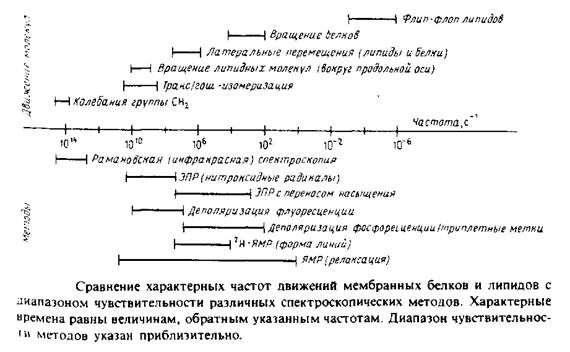
Диапазон движений, происходящих в мембране, весьма широк: от молекулярных колебаний с частотой порядка 1014 с до трансмембранного флип-флоп-переноса липидов, характерное время которого может достигать несколько суток. На рис. 1 в общем виде представлены некоторые из этих процессов, а также указаны временные пределы чувствительности различных биофизических методов. Величины, приведенные на рисунке, различаются на 20 порядков, поэтому термины «быстрый» и «медленный» не могут адекватно характеризовать различные типы движений. Из рисунка видно также, что одни методы позволяют получить статичную картину мембраны, поскольку характерное время соответствующих движений больше, чем время измерений, в то время как другие методы дают усредненную по времени картину, поскольку время перемещения молекул гораздо меньше, чем время измерения.
Мы рассмотрим два основных типа экспериментов. Первые основаны на использовании внутримембранных зондов для изучения текучести мембраны. Индикаторами физического состояния мембраны, а также характера липидно-белковых взаимодействий могут служить низкомолекулярные ЭПР-метки и флуоресцентные зонды.
Второй тип экспериментов направлен на прямое измерение латеральной диффузии мембранных белков или липидов и вращательной способности белков внутри бислоя. Исследовались также молекулярные взаимодействия в бислое, поскольку они влияют на динамику изучаемых молекул.
Другие статьи:
Концепция атомизма. Дискретность и непрерывность
материи
Строение материи интересует естествоиспытателей еще с античных времен. В Древней Греции обсуждались две противоположные гипотезы строения материальных тел. Одну из них предложил древнегреческий мыслитель Аристотель. Она заключается в том, ...
Выводы
Методы приспособления человеческой цивилизации к информационной революции и безопасности в информационной сфере могут быть заимствованы из нейрофизиологии и природы информационной системы самого человека.
Человечество в информационном пл ...
Охрана природы
Охрана природы – это система мер, направленных на поддержание рационального взаимодействия между человеком и окружающей природной средой, обеспечивающая сохранение и восстановление природных богатств, разумное использование природных ресу ...