Вывернутый белок

Относительно структуры бактериородопсина можно сделать весьма интересный вывод: его полипептидная цепь уложена так, что полярные или заряженные остатки, находящиеся в семи трансмембранных участках, оказываются обращенными внутрь глобулы, а неполярные контактируют с мембранными липидами. Глобула по существу "вывернута", если сравнивать ее с белками, в которых большинство неполярных остатков спрятаны внутри структуры, а полярные обращены наружу. Этому есть разумное объяснение, и об этом же свидетельствуют некоторые экспериментальные данные, в первую очередь данные по рассеянию нейтронов. В основе упомянутого метода лежит существенное различие между рассеянием на атомах водорода и дейтерия. Дейтерированные валин и фенилаланин включали путем биосинтеза в бактериородопсин и затем, используя разностный метод Фурье, устанавливали распределение этих аминокислот в проецируемой структуре пурпурной мембраны. Приняв общепринятые допущения о последовательности предполагаемых спиральных участков, при построении соответствующей модели обнаружили, что если смотреть вдоль оси спирали, то для каждой спирали остатки валина будут располагаться на сторонах, противоположных заряженным и полярным группам, а остатки фенилаланина - преимущественно на противоположной валину стороне. По данным нейтронного рассеяния, остатки валина преимущественно обращены к мембранным липидам, поэтому был сделан вывод, что заряженные и полярные остатки группируются внутри белка. Заметьте, что этот результат не зависит от конкретного расположения участков полипептида на электронно-микроскопической карте.
Другой подход основан на использовании гидрофобного фотореактивного зонда 1251-ТИД. Этот реагент внедряется в гидрофобную сердцевину мембраны и при фотолизе неспецифически реагирует с наиболее доступными боковыми группами аминокислот. Это его свойство может использоваться для локализации тех участков белковой молекулы, которые обращены к липидам. По крайней мере в двух случаях, в том числе в случае бактериородопсина, отмечалось включение метки в специфические, периодически расположенные участки полипептидной цепи, при этом метка включалась через каждые три или четыре остатка.
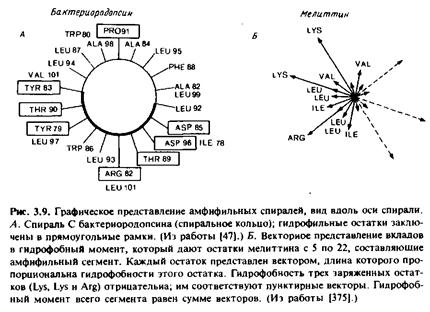
Если такой характер мечения отражает только долю боковых групп, обращенных к мембранным липидам, то мы сможем установить, какая сторона предполагаемого а-спирального участка контактирует с липидами. Поскольку на один виток спирали приходится 3,6 остатка, то наблюдаемая периодичность может иметь место только в том случае, если зонд контактирует лишь с одной стороной спирали. Данные по спирали С в бактериородопсине согласуются с представлением о вывернутой структуре этого белка и тоже свидетельствуют о том, что соответствующая часть полипептида имеет спиральную конфигурацию. В целом можно сказать, что концепция вывернутых интегральных мембранных белков весьма разумна, однако экспериментальное ее обоснование пока недостаточно. Напомним, что, согласно моделям фотосинтетического реакционного центра, в сердцевине бислоя находится очень мало ионизируемых остатков или они отсутствуют там вообще.
Другие статьи:
Конструирование рекомбинантных ДНК
Под рекомбинантными понимают ДНК, образованные объединением in vitro (в пробирке) двух или более фрагментов ДНК, выделенных из различных биологических источников. Ключевыми в этом определении являются слова "фрагмент ДНК" и &quo ...
Кальций
Наряду с фосфором кальций является основным минеральным компонентом костей и зубов. В них сосредоточено до 99% всего кальция организма. Кроме этого он участвует в процессах свертываемости крови, поддержании осмотического равновесия внутре ...
Были ли экологические кризисы, вызванные человеком, в древности? Опишите их
Слово "экология" чаще всего используют не в строгом значении, а в более узком, обозначая им взаимосвязи человека с окружающей средой, те изменения которые происходят благодаря антропогенному давлению в биосфере, равно как и проб ...