Механизмы сохранения
нуклеогидной последовательности ДНК. Химическая стабильность. Репликация. РепарацияСтраница 1

Для поддержания главных характеристик клетки или организма на протяжении их жизни, а также в ряду поколений наследственный материал должен отличаться устойчивостью к внешним воздействиям или должны существовать механизмы коррекции возникающих в нем изменений. В живой природе используются оба фактора. Третьим фактором является точность копирования нуклеотидных последовательностей материнской ДНК в процессе ее репликации.
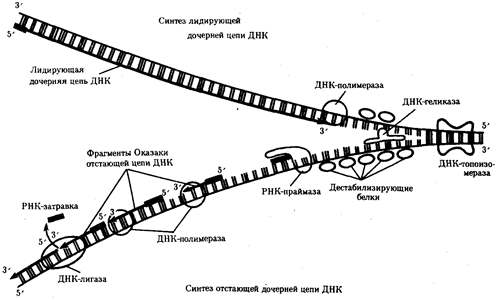
Рис.13. Белки, участвующие в процессе репликации ДНК
ДНК-геликаза расплетает двойную спираль ДНК, разделяя ее полинуклеотидные цепи; дестабилизирующие белки выпрямляют участок цепи ДНК; ДНК-топоизомераза разрывает фосфодиэфирную связь в одной из полинуглеотидных цепей ДНК, снимая напряжение, вызываемое расплетенисм спирали и расхождением цепей в репликационной вилке; РНК-праймаза синтезирует РНК-затравки для дочерней цепи и для каждого фрагмента Оказаки; ДНК-полимераза осуществляет непрерывный синтез лидирующей цепи и синтез фрагментов Оказаки отстающей цепи; ДНК-лигаза сшивает фрагменты Оказаки после удаления РНК-затравки
По реакционной способности молекулы ДНК относятся к категории химически инертных веществ. Известно, что роль вещества наследственности может выполнять не только ДНК, но и РНК (некоторые вирусы). Считают, что выбор в пользу ДНК обусловлен ее более низкой по сравнению с РНК реакционной способностью.
Рассмотренный выше механизм репликации отличается чрезвычайно высокой точностью воспроизведения структуры ДНК. При удвоении ДНК ошибки возникают в среднем с частотой 1·10-6 комплементарных пар оснований.
В поддержании высокой точности репликации важная роль принадлежит прежде всего ферменту ДНК-полимеразе. Этот фермент осуществляет отбор необходимых нуклеотидов из числа имеющихся в ядерном соке нуклеозидтрифосфатов (АТФ, ТТФ, ГТФ, ЦТФ), точное присоединение их к матричной цепи ДНК и включение в растущую дочернюю цепь. Частота включения неправильных нуклеотидов на этой стадии составляет 1·10-5 пар оснований.
Такие ошибки в работе ДНК-полимеразы связаны с возникновением измененных форм азотистых оснований, которые образуют "незаконные" пары с основаниями материнской цепи. Например, измененная форма цитозина вместо гуанина связывается водородными связями с аденином. В результате в растущую цепь ДНК включается ошибочный нуклеотид. Быстрый переход измененной формы такого основания в обычную нарушает его связывание с матрицей, появляется неспаренный 3'-ОН-конец растущей цепи ДНК. В этой ситуации включается механизм самокоррекции, осуществляемый ДНК-полимеразой (или тесно связанным с ней ферментом - редактирующей эндонуклеазой). Самокоррекция заключается в отщеплении ошибочно включенного в цепь ДНК нуклеотида, не спаренного с матрицей (рис.14). Следствием самокоррекции является снижение частоты ошибок в 10 раз (с 10-5 до 10-6).
Несмотря на эффективность самокоррекции, в ходе репликации после удвоения ДНК в ней обнаруживаются ошибки. Особенно часто это наблюдается при нарушении концентрации четырех нуклеозидтрифосфатов в окружающем субстрате. Значительная часть изменений возникает также в молекулах ДНК в результате спонтанно происходящих процессов, связанных с потерей пуриновых оснований - аденина и гуанина (апуринизацией) - или дезаминированием цитозина, который превращается в урацил. Частота последних изменений достигает 100 на 1 геном/сут.
Содержащиеся в ДНК основания могут изменяться под влиянием реакционноспособных соединений, нарушающих их нормальное спаривание, а также под действием ультрафиолетового излучения, которое может вызвать образование ковалентной связи между двумя соседними остатками тимина в ДНК (димеры тимина). Названные изменения в очередном цикле репликации должны привести либо к выпадению пар оснований в дочерней ДНК, либо к замене одних пар другими. Указанные изменения действительно сопровождают каждый цикл репликации ДНК, однако их частота значительно меньше, чем должна была бы быть. Это объясняется тем, что большинство изменений такого рода устраняется благодаря действию механизма репарации (молекулярного восстановления) исходной нуклеотидной последовательности ДНК.
Другие статьи:
Характеристика семейства Лютиковые. Нарисуйте разные типы цветков и плодов.
Формулы цветков. Укажите представителей 15 видов и их практическое значение
Лютиковые преимущественно травянистые растения, иногда лианы. Листья очередные, реже супротивные. Цветки (у первичных родов примитивного типа) правильные и неправильные, спирально-циклические, одиночные или в малоцветковых соцветиях. Опыл ...
Обучение безопасности труда и виды инструктажа
Обучение безопасным приёмам труда для работников проводится на основании государственного стандарта – ТОСТ 12.0.004–90 ССБТ. Организация обучения по безопасности труда. Общие положения». Необходимость обучения и инструктирования работнико ...
Определение способности плесневых грибов
использовать соединения углерода
Микромицеты характеризуются неодинаковой способностью использовать различные соединения углерода для конструктивного и энергетического метаболизма. Чтобы выяснить возможность роста гриба за счет тех или иных углеродсодержащих веществ, их ...