Жидкие кристаллыСтраница 1

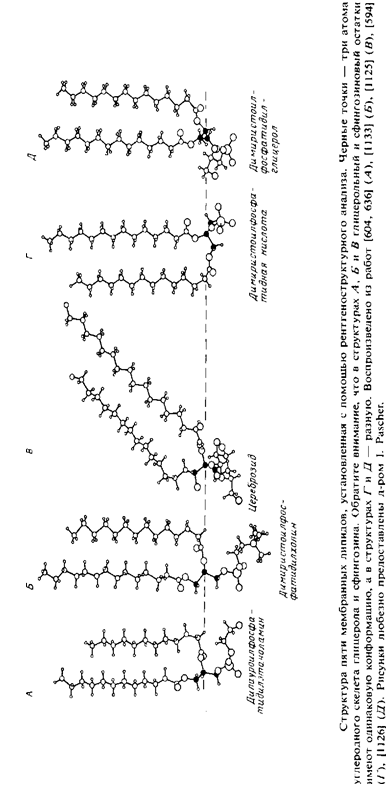
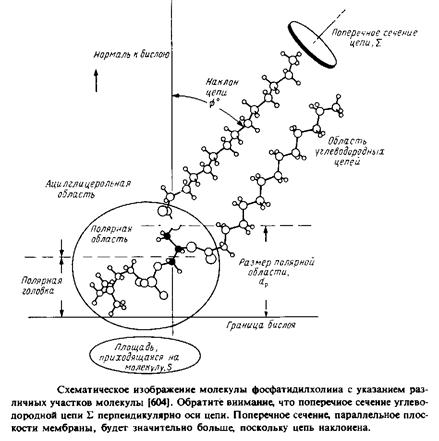
С помощью ренгтеноструктурного анализа установлена с высоким разрешением пространственная структура ряда мембранных липидов. К ним относится лизофосфатидилхолин, димиристо-илфосфатидная кислота, димиристоилфосфатидилхолин, дилауроилфосфатидилэтаноламин, димиристоилфосфатилглицерол и цереброзид. Кристаллы этих липидов содержат очень мало воды, однако пространственная структура липидов в них оказалась подобна той, которую они имеют в полностью ги-дратированном состоянии. На рис. представлена пространственная структура некоторых из этих мембранных липидов в кристаллах. Атомы углерода остатка глицерола и соответствующие атомы Сфингозина выделены черным цветом. На рис. указаны некоторые структурные параметры, используемые для описания конфор-мации липидов.
Рассмотрим кристаллическую структуру дилауроилфосфатидил-этаноамина и укажем наиболее характерные ее особенности.
1. Площадь, приходящаяся на молекулу, составляет 39 А2.
2. Полярная головка в целом ориентирована параллельно плоскости бислоя. При этом аминогруппа образует водородную связь с неэтерифицированными атомами кислорода фосфатной группы соседней молекулы. Остаток глицерола ориентирован перпендикулярно плоскости бислоя.
3. Жирнокислотная цепь sn-2 сначала идет параллельно поверхности бислоя, а после второго углеродного атома направляется в глубь бислоя.
4. Ацильные цепи расположены перпендикулярно поверхности бислоя и, за исключением начального участка sn-2 жирнокислотной цепи, максимально вытянуты, т. е. имеют полностью-ти/имс-конфигурацию.
Кристаллические структуры фосфатидилхолина и цереброзида во многом сходны со структурой фосфаридилэтаноламина, хотя и
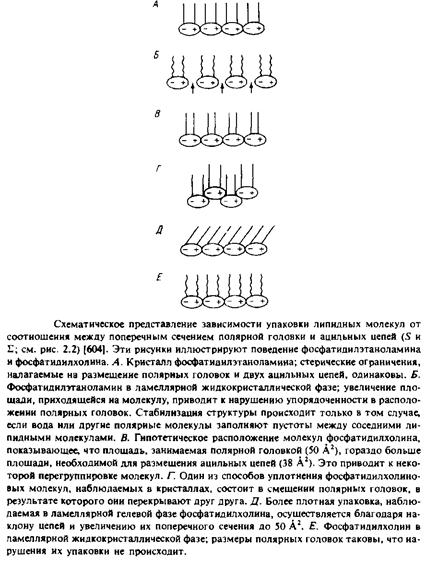
имеют ряд важных отличий. Наиболее значительное и явное из них состоит в наклоне ацильных цепей, сильно выраженном в случае цереброзида. Этот наклон связан с наличием стерических препятствий для упаковки молекул. Объемные полярные головки фосфати-дилхолина и цереброзида не позволяют им упаковываться в сравнительно простые структуры, как в случае дилауроилфосфатидилэта-ноламина. Площадь S, необходимая для размещения этих головок, превышает величину 39 А2, приходящуюся на поперечное сечение ацильных цепей каждой молекулы. В случае цереброзида эта проблема решается за счет наклона ацильных цепей по отношению к плоскости бислоя. В результате существенно увеличивается площадь проекции поперечного сечения ацильных цепей на плоскость бислоя. На рис. схематично показано, как благодаря наклону цепей сохраняется взаимодействие между цепями соседних молекул и обеспечивается размещение объемных полярных групп. Жирнокислотные цепи димиристоилфосфатидилхолина отклоняются от нормали к поверхности бислоя всего на 12°, тогда как в случае цереброзида это отклонение достигает 41°. Проблема упаковки объемных полярных групп диацилфосфатидилхолина может быть решена путем поочередного смещения соседних молекул вдоль нормали к бислою, как схематически показано на рис. Л Имеются убедительные данные и о значительном наклоне цепей в гелевой фазе полностью гидратированных липидных бислоев. Это наглядный пример того, как простые стерические соображения, учитывающие «форму» липидных молекул, оказываются весьма полезными при рассмотрении пространственной структуры липидов.
В кристаллах всех изученных липидов, за исключением фосфа-тидной кислоты, начальный участок 5«-2-жирнокислотной цепи направлен параллельно поверхности бислоя. Об этой особенности расположения цепей свидетельствовали также результаты ЯМР-иссле-дований фосфатидилэтаноламиновых и фосфатидилхолиновых бислоев, а также фосфолипидов в мембранах Е. coli. Физиологическая значимость этой структурной особенности неясна. Однако отмечалось, что в яичном фосфатидилхолине средняя длина sn-2-жирнокислотной цепи равна 18 углеродным атомам, а средняя длина sn-1-цепи — 16. По-видимому, это позволяет скомпенсировать излом жирнокислотной цепи во втором положении остатка глице-рола так, что обе ацильные цепи оказываются погруженными в бислой на одну и ту же глубину.
Другие статьи:
Биологический ритмы и возраст
Онтогенные аспекты биоритмологии имеют важное значение для профилактики нарушений здоровья, что нашло отражение в литературе.
В.А. Доскин и Н.Н. Куинджи обобщили современные сведения о биоритмах у детей и подростков. Авторы преимуществен ...
Ритмы космоса и здоровье человека
Биологические ритмы — это один из механизмов, которые позволяют организму приспосабливаться к меняющимся условиям жизни. Подобная адаптация происходит в течение всей нашей жизни, ибо постоянно происходит и изменение внешней среды. Сменяют ...
Описание рода. Род спирея или таволга (Spiraea L.). Сем.
Розоцветные (Rosaceae Juss.)
В роде около 90 видов. В СНГ дико произрастают 25, интродуцировано 66. Это невысокие листопадные кустарники 0,5 – 3,0 м высоты, с пышными раскидистыми кронами, изящными тонкими побегами и обильными соцветиями. Цветут в различное время. Бе ...