Сегментация и деление тела на отделыСтраница 1

Тело многощетинковых червей поделено на сегменты (членики), которые внешне хорошо распознаются по наличию на каждом из них по одной паре параподий, снабженных обычно различными придатками. Наружная кольчатость тела тесно связана с внутренними септами (диссепиментами),: обособляющими друг от друга отдельные сегменты (метамеры, или сомиты) (рис. 1). В каждом сегменте, как правило, закономерно повторяются наборы внутренних органов; комплекс нервных образований одного сегмента обозначается как невросомит, сплетение кровеносных сосудов — ангиосомит, система мышечных волокон — миосомит и т. д.; большей частью метамерно располагаются также выделительные органы и гонады. У чешуйчатых червей кишечник в каждом сегменте имеет боковые слепые выросты (карманы, или дивертикулы), так что и он у них обладает вполне определенной метамерией (рис. 2).
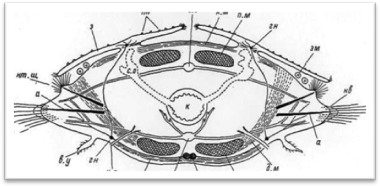
Рис. 1. Поперечный разрез через туловищный сомит (чешуйчатый сегмент).
Таким образом, наружная кольчатость, или наружная метамерия, червей обычно соответствует внутренней метамерии.1
Полная гомономность (однородность) всех туловищных сегментов у многощетинковых червей никогда не наблюдается. Наибольшая гомономность сегментов характерна для бродячих (эррантных) полихет, но и у них она в той или другой степени нарушена. Следует различать гетерономность первичную и вторичную. Первичная гетерономность возникает еще в процессе метаморфоза личинки и связана с образованием ларвальных и постларвальных сегментов, имеющих разное происхождение. Ларвальные сегменты закладываются обычно одновременно на начальной стадии метаморфоза личинок, а лостларвальные сегменты последовательно и постепенно возникают из предпигидиальной эктодермальной зоны роста, причем целом в ларвальных сегментах всегда формируется из мезодермальных полосок путем их разделения на мезодермальные блоки, которые затем становятся полыми.
1 При описании и на рисунках червей сегменты обозначаются римскими цифрами
Число ларвальных сегментов варьирует: у Polynoidae их 8—10, а у Siga-lionidae — 4—6. Большое и изменчивое количество ларвальных сегментов указывает на примитивный тип метаморфоза. Ларвальные сегменты в отличие от постларвальных не образуют половых желез, не имеют целомодуктов и хлора-гогенных клеток. Иногда ларвальные сегменты подвергаются слиянию, образуя перистомиум: например, крупный перистомиум нереид представляет собой продукт слияния трех передних ларвальных сегментов. Вторичная гетерономность относится к постларвальным сегментам, она целиком обусловлена особенностями обитания взрослых червей (адаптивные приспособления).
Количество сегментов у чешуйчатых червей, как и у большинства других полихет, сильно варьирует. Существуют виды, у которых сегментов относительно немного и их число строго постоянное (например, Lepidonotus — всего 26 сегментов), но некоторые виды (многие Lepidasthenia, Arctonoe и др.) состоят из 100 и более сегментов, причем количество последних увеличивается с возрастом и возникновение новых сегментов на заднем конце тела (в предпиги-диальной зоне роста) не прекращается с наступлением половой зрелости. Самые задние сегменты обычно меньше (тоньше и короче) остальных и имеют недоразвитые параподий. Вполне естественно, что черви с большим количеством сегментов одновременно и наиболее длинные. Однако, как уже отмечалось ранее, количество сегментов не всегда строго коррелирует с размерами самих червей. Неопределенно большое количество сегментов у полиноидных червей мы рассматриваем как примитивное состояние, а стабилизацию количества сегментов — как прогрессивный момент. Подсчет сегментов легче всего производить с брюшной стороны, так как спинная сторона червей нередко замаскирована различными придатками, в частности у чешуйчатых червей элитрами. Количество сегментов у представителей афродитид (Aphro-f dita, Laetmonice, Hermonia, Pontogenia) не превышает 50 (колеблется в пределах 32—47). Среди полипоид по числу сегментов выделяется три группы родов: 1) сегментов 25— 30 — Lepidonotus, Hermenia, Euphione, Iphione; 2) сегментов 30—50 — Harmothoe, Lagisca, Eunoe, Antinoe, Gattyana, Bylgides, Eucranta, Malm-grenia, Alentia и др. и 3) сегментов свыше 50 (до 100 и более) — Perolepis, Arctonoe, Lepidasthenia и др. Для большинства сигалионид характерно более 50 сегментов.
Другие статьи:
Приготовление растворов
1. 0,5 % раствор крахмала. В дистиллированную воду объёмом 30 мл прибавили 0,5 г порошка крахмала, тщательно перемешали и нагрели до кипения. В полученный раствор прилили 70 мл холодной дистиллированной воды и охладили.
2. Раствор 0,003 ...
Практическая часть
Анкета
1)
Испытываешь ли ты чувство дискомфорта в области желудка после еды?
2)
Имеете ли вы какие-нибудь врожденные болезни?
3)
Здоровые ли у вас зубы?
4)
Употребляете ли вы фрукты во время еды?
5)
Часто ли вы едите однообразну ...
Сценарии
будущего человека. Старение как естественный процесс
Старение и смерть любого живого организма, в том числе и человека, — естественный процесс. Именно так диалектика жизни и смерть трактовалась в системе прошлых культур.
С самых древних времен и до XVIII в. считалось, что старение обусловл ...