Результаты и обсуждение.Страница 2

Экспериментально разработанный способ получения дейтерий-меченного инозина представлен на схеме. Его основные этапы: наработка дейтерированной биомассы метилотрофных бактерий B. methylicum путем культивирования бактерий в полностью дейтерированной среде dM9; выделение фракции общих белков биомассы и их гидролиз; ферментация базового штамма B. subtilis на среде, приготовленной на основе 99,9 ат% 2Н2О и гидролизатов биомассы метилотрофных бактерий; выделение инозина, включая его адсорбцию активированным углем, десорбцию спиртово-аммиачным (1:1) раствором и последующую кристаллизацию инозина из метанола. Чистоту полученного инозина контролировали методом тонкослойной хроматографии (ТСХ), используя в качестве сравнения хроматографически чистые стандарты нуклеозидов. ТСХ инозина, выделенного из культуральной жидкости, показала наличие в анализируемом образце единственного пятна с Rf = 0,51, соответствующего по подвижности чистому инозину.
Изучение ростовых и биосинтетических характеристик B. subtilis
. При изучении роста штамма B. subtillis и уровня накопления инозина в КЖ использовали следующие среды:
1). Ферментационная среда (FM-среда), приготовленная стандартно на обычной воде.
2). Ферментационная среда, приготовленная из 99,9 ат.% 2Н2O (dFM-среда) и содержащяя дейтерий-меченную биомассу метилотрофных бактерий, выделенную, соответственно из среды dМ9.
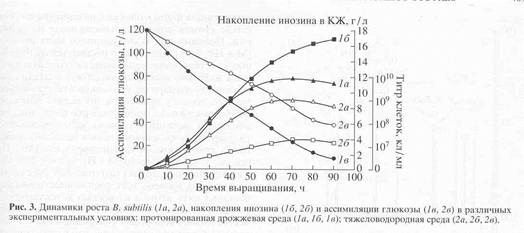
Кривые, отражающие динамику роста, ассимиляции глюкозы и накопление инозина в культуральной жидкости штаммом B. subtilis в условиях протонированной среды и среды, содержащей тяжелую воду и гидролизаты биомассы метилотрофных бактерий, представлены на рис.3. Сравнивая данные по росту штамма на протонированной и dFM-среде, можно заключить, что рост B. subtilis на dFM-среде (рис.3) слабо ингибируется дейтерием, поэтому выход биомассы, продолжительность лаг-фазы и длительность времени клеточной генерации при переносе клеток B. subtillis со стандартной на дейтерированную среду в целом изменяются незначительно. Как видно из рис.3, при росте штамма на среде, содержащей обычную воду уровень накопления инозина в культуральной жидкости достигал величины 18,3 г/л после пяти суток культивирования. Вместе с тем, биосинтез инозина на dFМ-среде, был снижен в 4,6 раз по-сравнению с исходным штаммом на протонированной среде (рис.3). Такие низкие уровни секреции инозина на dFM-cреде коррелируют со степенью конверсии глюкозы в этих условиях. Как видно из рис.3, кривая конверсии глюкозы на полностью дейтерированной среде имеет меньший угол наклона, чем на среде с обычной водой, что свидетельствует о том, что при росте на 2Н2O глюкоза расходуется менее эффективно. Вместе с тем мы не исключаем, что все эти вышеназванные эффекты могут возникать не вследствие ингибирования дейтерием биосинтеза , а в результате неэквивалентной замены БВК на биомассу метилотрофных бактерий. Предметом дальнейших исследований будет разработка и оптимизация сбалансированных по ростовым факторам сред на основе биомассы метилотрофных бактерий.
Полученные данные в целом подтверждают устойчивое представление о том, что адаптация к 2Н2О является фенотипическим явлением, поскольку адаптированные к тяжелой воде клетки возвращаются к нормальному росту и биосинтезу в протонированных средах после некоторого лаг-периода. В то же время обратимость роста на 2Н2O и Н2O средах теоретически не исключает возможности того, что этот признак стабильно сохраняется при росте в Н2О, но маскируется при переносе клеток на дейтерированную среду. Можно предположить, что клетка реализует лабильные адаптивные механизмы, которые способствуют функциональной реорганизации работы жизненно-важных систем в 2Н2O. Нам представляется выбор бактерий в качестве модельных объектов для этих целей наиболее целесообразным, так как прокариоты как организмы, стоящие на более низких ступенях развития живого,наиболее лабильны в генетическом аспекте и тем самым быстрее реагируют и приспосабливаются к изменчивым факторам среды. Для того чтобы сделать более конкретные выводы о природе и механизме адаптации клеток к тяжелой воде, необходимы экспериментальные данные по физиологии и биохимии адаптированных клеток.
Другие статьи:
Гаметогенез и развитие растений
У растений гаметогенез протекает значительно сложнее. При этом процесс мейоза имеет место не на стадии образования гамет, а на стадии образования спор. Кроме того, у растений наблюдается чередование поколений с диплоидным и гаплоидным наб ...
Транспорт глюкозы.
Транспорт глюкозы может происходить по типу как облегченной диффузии, так и активного транспорта, причем в первом случае он протекает как унипорт, во втором – как симпорт. Глюкоза может транспортироваться в эритроциты путем облегченной ди ...
sTNF-RII
Растворимый рецептор интерлейкина-2 ...