Структура фотосинтетических реакционных центров ft. viridis и ft.
sphaeroidesСтраница 1

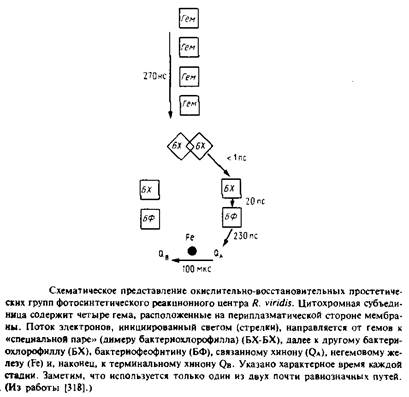
Фотосинтетические реакционные центры представляют собой комплексы белков с пигментами; в них происходит первичное разделение зарядов в фотосинтетических мембранах. Лучше всего охарактеризованы комплексы из пурпурных несерных бактерий; они обычно состоят из трех белковых субъединиц - Н, М и L. Реакционный центр Rhodopseudomonas viridis имеет также четвертую субъединицу - цитохром с-типа. Простетическими группами этого комплекса являются четыре гемогруппы, четыре бактериохлорофилла Ь, два бактериофеофитина, одно негемовое железо, один менахинон и один убихинон. Под действием света электрон переходит от первичного донора электронов, так называемой "специальной пары" - молекул бактериохлорофилла, образующих димер, к бактериофеофитину, а затем к первичному хиноновому акцептору QA. В конце концов электрон восстанавливает вторичный акцептор Qb в ходе реакции, при которой протоны поступают из раствора на восстановленный хинон. Qb находится в равновесии с хинонным пулом в би-слое. Окисленный первичный донор электронов, "специальная пара", восстанавливается цитохромом с-типа. Поскольку цитохром и хинон расположены на противоположных сторонах фотосинтетической мембраны, светозависимый электронный транспорт электроге-нен и генерирует трансмембранную разность потенциалов.
Суммарная мол. масса реакционного центра из R. viridis составляет примерно 150 ООО, а кажущаяся мол. масса субъединиц - 38 ООО, 35 ООО, 28 ООО и 24 ООО. Заметим, что электрофорез в ПААГ-ДСН дает неправильные молекулярные массы для Н, М и L субъединиц; об этом свидетельствуют данные о числе аминокислотных остатков в каждом полипептиде, полученные при секвенировании ДНК. Очищенный комплекс был закристаллизован с использованием сульфата аммония как осаждающего агента в присутствии детергента N. N-диме-тилдодециламин-М-оксида и органического амфифильного соединения гептан-1,2,3-триола. Кристаллы были в достаточной степени упорядочены и фотохимически активны. Структура этого четырехсубъединичного белка была установлена с разрешением около 3 А. Детергент в кристаллах не упорядочен, поэтому невозможно точно определить положения границ погруженных в мембрану участков. Размеры комплекса - 30 х 70 х 130 A. L - и М-субъединицы содержат по пять трансмембранных а-спиральных участков, а у Н-субъединицы такой участок только один.
Итак, все трансмембранные области этого белкового комплекса имеют а-спиральную конфигурацию. Длина каждого а-спирального сегмента составляет примерно 40 А, этого достаточно для пересечения мембраны.
Структура белкового комплекса из R. viridis напоминает сэндвич. L - и М-субъединицы уложены одинаковым образом и расположены в центре сэндвича. Они пересекают бислой и связаны со всеми простетическими группами, за исключением гемов. Сегменты L и М, которые соединяют трансмембранные сегменты по обе стороны мембраны, участвуют в связывании цитохрома и Н-субъединиц. Цитохром образует "шапочку" на наружной поверхности бислоя, а гидрофильная часть субъединицы Н - аналогичную структуру на цитоплазматической поверхности. Трансмембранная а-спираль на N-конце Н-субъединицы контактирует с цитохромом на противоположной стороне мембраны.
Другие статьи:
Организация
биосферы
С появлением динозавров резко усложнялась организация биосферы, выразившаяся в образовании различных экосистем со сложной и разнообразной структурой и множеством трофических цепей, в кайнозое, при господстве млекопитающих, биосфера повтор ...
Гибридные спирали ДНК-РНК
Полинуклеотидные цепочки РНК и ДНК, имеющие комплементарные последовательности, могут образовывать двойную спираль РНК-ДНК с антипараллельными цепями. Структура таких комплексов напоминает А-ДНК. Спаривание оснований в них отвечает правил ...
Реакционная способность веществ, анализ и синтез. Реакционная
способность веществ
Реакционная способность, характеристика относительной хим. активности молекул, атомов, ионов, радикалов. Для количественной оценки рассматривают реакционные серии, т.е. ряды однотипных реакций, проводимых в одинаковых условиях, например: ...