Особенности организации
и экспрессии генетической информации у про- и
эукариотСтраница 3

Возможно, область терминатора, богатая Г-Ц парами, играет определенную роль в остановке РНК-полимеразы, а участок РНК, содержащий УУУУ обеспечивает отделение транскрипта от матрицы ДНК.
У эукариот не обнаружено образования структур, подобных шпилькам в прокариотических РНК. Поэтому, каким образом у них осуществляется терминация транскрипции, остается неясным. Cbd oil uk
В составе всех мРНК можно выделить кодирующие участки, представляющие набор кодонов, которые шифруют последовательность аминокислот в пептиде. Как правило, эти участки начинаются стартовым кодоном АУГ, но иногда у бактерий используется кодон ГУТ. На конце кодирующей последовательности располагается терминирующий кодон. Помимо кодирующих участков в мРНК на обоих концах могут располагаться дополнительные последовательности. На 5'-конце это лидерный участок, расположенный перед стартовым кодоном. На 3'-конце - трейлер, следующий за кодоном-терминатором.
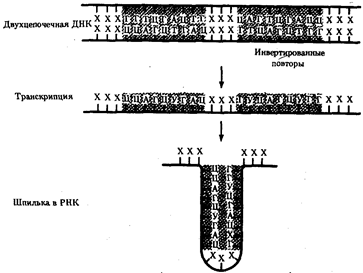
Рис.38. Образование шпильки участком РНК при терминации транскрипции у прокариот
Область РНК, несущая палиндром, образует комплементарно спаривающуюся структуру - шпильку (инвертированные повторы заштрихованы)
В полицистронной мРНК прокариот между кодирующими участками имеются межцистронные области, варьирующие по размерам (рис.3.39).
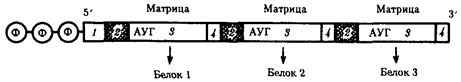
Рис.39. Полицистронная матричная РНК прокариот:
1 - некодирующие области, 2 - межцистронные области, 3 - кодирующие области, 4 - терминирующие кодоны
В связи с тем что прокариотические гены целиком состоят из нуклеотидных последовательностей, участвующих в кодировании информации, транскрибированные с них РНК сразу после их синтеза способны выполнять функцию матриц для трансляции. Лишь в исключительных случаях требуется их предварительное созревание - процессинг.
В отличие от прокариотических генов большинство генов эукариотических клеток прерывисты, так как несут в своем составе неинформативные нуклеотидные последовательности - интроны, не участвующие в кодировании информации. В связи с этим первичные транскрипты, синтезированные РНК-полимеразой II, обладают большими, чем необходимо для трансляции, размерами и оказываются менее стабильными. В совокупности они образуют так называемую гетерогенную ядерную РНК (тРНК), которая прежде чем выйти из ядра и начать активно функционировать в цитоплазме, подвергается процессингу и превращается в зрелые мРНК.
Процессинг эукариотических мРНК. Созревание, или процессинг, мРНК предполагает модифицирование первичного транскрипта и удаление из него некодирующих интронных участков с последующим соединением (сплайсингом) кодирующих последовательностей - экзонов. Модифицирование первичного транскрипта эукариотической мРНК начинается вскоре после синтеза его 5'-конца, содержащего одно из пуриновых оснований (аденин или гуанин). На этом конце образуется колпачок - кэп, который блокирует 5'-конец мРНК путем присоединения к первому нуклеотиду транскрипта трифосфонуклеозида, содержащего гуанин, связью 5'-5'.
Гффф + фффАфN… → ГфффАфN. + фф + ф В результате образуется последовательность ГфффАфЧМ ., в которой остаток туанина находится в обратной ориентации по отношению к другим нуклеотидам мРНК. Модификация 5'-конца мРНК предполагает также метилирование присоединенного гуанина и первых двух-трех оснований первичного транскрипта (рис.3.40). Образуемые на 5' - концах мРНК кэпы обеспечивают узнавание молекул мРНК малыми субчастицами рибосом в цитоплазме. Кэширование осуществляется еще до окончания синтеза первичного транскрипта.
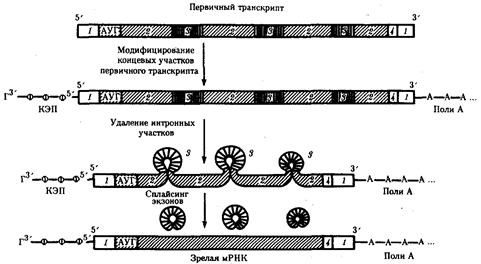
Рис. 40. Образование зрелой мРНК эукариот в ходе процессинга:
1 - некодирующие последовательности, 2 - экзоны, 3 - интроны, 4 - кодон-терминатор
После завершения транскрипции происходит удаление части нуклеотидов на 3'-конце первичного транскрипта и присоединение к нему последовательности, состоящей из 100-200 остатков адениловой кислоты (полиА) (рис.3.40). Считают, что эта последовательность способствует дальнейшему процессингу и транспорту зрелой мРНК из ядра. После выхода мРНК в цитоплазму ее полиА-последовательность постепенно укорачивается под действием ферментов, отщепляющих нуклеотиды на 3'-конце. Таким образом, по длине полиА-последовательности можно косвенно судить о времени пребывания мРНК в цитоплазме. Возможно, добавление полиА-последовательности в ходе процессинга повышает стабильность мРНК. Однако около трети мРНК вообще не содержат полиА-участка. К ним относятся, например, гистоновые мРНК.
Другие статьи:
Программа “геном человека”
Геном секвенировали в 2003 г‚ т.е. к пятидесятилетнему юбилею открытия двойной спирали ДНК (1953)‚ планировалось к 2005 г.
В 1988 г. один из первооткрывателей знаменитой двойной спирали ДНК, нобелевский лауреат Дж. Уотсон, публично выска ...
Результаты исследований
В ходе курсовой работы изучили влияние таких антибиотиков, как стрептомицин, пенициллин, ампициллин, на грамположительные бактерии рода Bacillus: Bacillus subtilis, Bacillus mesentericus, Bacillus cereus.
Bacillus cereus образовал выпукл ...
Пища, приготовленная природой
Молоко— идеальный продукт питания. Павлов писал, что среди других видов человеческой еды особое место отведено молоку. Это «пища, приготовленная самой природой». Она входит в рацион всех цивилизованных народов с древнейших времен.
Археол ...